Inheritance: Genes at Work: Mendel's Breakthrough
Genes at Work: Mendel's Breakthrough
Hippocrates hypothesized that particles called pangenes occur in all parts of the body, and change as a person matures to reflect body changes, and then enter the egg or sperm of an individual to be passed on to the next generation. Although this idea seemed to make sense and was widely held as correct at the time, it unfortunately did not account for genetic abnormalities or even common human discrepancies such as hair- or eye-color variations.
Pangenesis was replaced in the early 1800s by the hypothesis of “blending,” which stated that offspring inherit and blend traits from their parents. For instance, a red flower crossed with a white flower should always produce a pink flower. Unfortunately, this is not always the case; sometimes a red flower and a white flower produce a red flower or a white flower, not necessarily pink.
A student at the University of Vienna published a paper in 1866 that became the foundation for modern genetics. Gregor Mendel, an Augustinian monk, grew and investigated garden peas and subsequently explained “heritable” factors. He also posed and confirmed several salient hypotheses and instituted a new vocabulary to explain his findings that is still in use today.
There is more than one form for a gene (allele). For example, based on his experimentation with purple and white flowering pea plants, he deduced after examining the results of numerous crossings, that one gene is responsible for the purple color and a different gene is responsible for the white color of the flowers.
In most cases, two genes are needed to express a trait, one from each parent. Likewise, if an organism is true-breeding, both genes that control the same trait had to be the same gene (homozygous).
The principle of segregation states that during egg and sperm formation, the pair of genes separates (segregates) and only one gene becomes incorporated into either the sperm or the egg, and the other gene can also become incorporated into either a sperm or egg, so that when sperm and egg unite, the offspring has the normal number, or pair, of genes.
When two different alleles for the same trait are found in an organism, the gene that is expressed is the dominant allele, and the gene not expressed is the recessive allele. If red is dominant over white in flowers, the offspring between a homozygous red and white flower should all be red.
Bionote
A Punnet square is used to predict the possible genotype and phenotype of offspring. It operates on Mendelean principles. As a reference, the genotype is the genetic makeup of an individual and the phenotype is the expression of the genetic makeup. In other words, the phenotype is what the organism looks like.
The principle of independent assortment states that each gene from the pair of alleles separates independently during gamete formation. Mendel noticed that during a dihybrid cross (cross of two traits), the offspring inherited the alleles as if they separated from each other and had equal chance of inheritance as the alleles from the other trait. Specifically, Mendel crossed a plant that was homozygous dominant for round, yellow seeds (RRYY, R = round, Y = yellow) with a homozygous plant with wrinkled, green seeds (rryy, r = wrinkled, y = green). If the genes stuck together and did not act independently, he hypothesized that the F2 generation must contain the same genotype as the parents, either RY or ry. For reference purposes, the F1 generation is the product of the mating of the original parents; the F2 is the product of the F2 mating. The F2 then becomes the “grandchildren” of the original mating. Through experimentation he found the following genotypes: RY, Ry, rY, ry. He correctly concluded that the individual alleles were segregating from the paired alleles and were free to unite in all possible combinations. Refer to the illustration F1 Punnet square. It can be used to predict the results of an RY and ry mating.
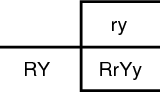
First, the genotype of the parents must be known. In this case they are RRYY and rryy. Next they are segregated. If they independently sort, the possible genotypes are RY and ry.
All of the F1 (first generation) offspring are heterozygous, meaning they contained both a dominant and recessive gene, RrYy, round and yellow. However, in the F2 generation, interesting data revealed a variety of offspring. Refer to the illustration F2 Punnet Square. Parent's genotype: RrYy ¥ RrYy.

Offspring phenotype:
- Round yellow = 9, round green = 3
- Wrinkled yellow = 3, wrinkled green = 1
The gametes had to sort independent of each other as depicted. The phenotypic ratio of 9:3:3:1 is typical of a dihybrid cross.
Mendel could tell by the phenotypes of the offspring that the gametes had recombined in such a way that they had to have an equal chance of combining with the other allele.
A test cross determines an unknown genotype by mating it with a homozygous recessive. This works for two reasons: First, the homozygous organism is an obvious true breeder. Second, the proof is in the offspring; if a homozygous recessive crosses with a homozygous dominant, all offspring are dominant; if matched with a heterozygous mate, some of the offspring will exhibit dominant and other recessive traits; a cross with another homozygous recessive produces only homozygous recessive offspring.
Test Cross

The following are the three possible test crosses that can occur. The researcher simply has to examine the phenotype to determine the unknown genotype of one of the parents. Let's simplify and only consider one gene: G is dominant for green; g is recessive for white. In every test cross, the unknown parent is always mated with the homozygous recessive parent. In this case, the test-cross parent's genotype is gg. There are three possible genotypes for the unknown parent: GG, Gg, and gg. Consider the following matings:

- In Case 1, all of the offspring are green. So when a test cross gives offspring that exhibit the dominant phenotype, the unknown parent must have been homozygous dominant. Refer to the illustration Case 1 (all green).
- In Case 2, half of the offspring exhibit the dominant phenotype, and the other half exhibits the recessive phenotype. When the offspring of a test cross exhibit a 50:50 ratio of dominant to recessive, the parent had to be heterozygous. Refer to the illustration Case 2 (half green, half white).

- In Case 3, all offspring are white. The only way this could happen is if the unknown parent was homozygous recessive. Refer to the illustration Case 3 (all white).
A test cross is an easy way to experimentally determine the unknown genotype of a parent. But remember, the genotype of the other parent must be homozygous recessive.
Pedigree
Pedigrees are useful in tracking traits through ancestors and for predicting traits for descendents. Refer to the illustration Pedigree.
Assume that a “**” behind the name indicates homozygous dominant for a fictitious trait. “*-” is heterozygous, and “--” is homozygous recessive for that trait.

The male and female mate and produce two sons, numbers 4 and 7, and a daughter, number 6 in the F1 generation. They each mate and produce the offspring shown in F2. An analysis of this pedigree shows that the fictitious trait is inherited by normal genetic inheritance patterns. For instance, the gene is not linked to the X chromosome (see Genetic Variation and Natural Selection).
Pedigrees for pets are common to ensure that the new puppy will grow into the type of dog the owner intended to purchase. Human pedigrees are also common, but in a different way. Genetic counseling provides potential parents with the likelihood that their children will inherit a specific disease. Also, most medical doctors ask as a routine question whether a history of heart disease or cancer exists in your family tree. Both are examples of the usefulness of a family pedigree.

Excerpted from The Complete Idiot's Guide to Biology © 2004 by Glen E. Moulton, Ed.D.. All rights reserved including the right of reproduction in whole or in part in any form. Used by arrangement with Alpha Books, a member of Penguin Group (USA) Inc.
To order this book direct from the publisher, visit the Penguin USA website or call 1-800-253-6476. You can also purchase this book at Amazon.com and Barnes & Noble.