Anatomy and Physiology: You've Got Potential
You've Got Potential
If you remember the basics of polarized membranes from The Structure of the Muscles and Muscle Cells, this will all be familiar. An inactive neuron is not truly at rest, for it requires a membrane to have an uneven concentration of ions on both sides of the neurilemma (the nerve cell membrane); a membrane in this state is said to be polarized. Ions and molecules naturally move to a state of equilibrium through a form of passive transport called diffusion. To be uneven, active transport is needed, which requires energy in the form of ATP.
Nerve impulses require some form of stimulus to initiate the depolarization of the membrane. Since that depolarization is accomplished through facilitated diffusion, in which ion channels open in the membrane, causing a rapid movement of ions, this combination of active transport and facilitated diffusion helps explain the rapidity of our response.
There are many types of ion channels which will initiate the depolarization of the membrane. All of these channels are gated channels, which require some environmental change to open them. These are opposed to leakage or nongated channels, which are so named because they are always open. A chemically gated ion channel is typically found at synapses, such as the cholinergic receptors that open in the presence of ACh (in both muscle cells and neurons). Such chemical channels open directly as a result of the neurotransmitter, for the receptor becomes a channel protein when the neurotransmitter binds to it. Indirect chemically gated channels function very similar to the G-protein/secondary messenger systems involved with some hormones.
Light-gated ion channels open, as the name implies, to the presence of light, which is why they are found in the retina. Mechanically gated ion channels, on the other hand, open in response to pressure, or vibration, which explains their presence in the ear (for both sound and for balance), and in the skin for various forms of touch. The Senses explores the various senses much further.
The last to consider is called the voltage-gated ion channel, which is essential to the basic conduction of nerve impulses. Its name refers to the current produced by the movement of ions, rather than the movement of electrons as in electricity. As a result of these channels, the depolarization of the membrane in one area results in the depolarization of a neighboring channel, and so on, ultimately producing an impulse. This makes cells with these channels (muscle cells and neurons) excitable, which means they are capable of producing an impulse, otherwise known as a traveling wave of depolarization and repolarization.
All for One? All or None!
Throughout this section you might have noticed the similarities between muscle and nerve cells. The same polarization, the same depolarization, the same neurotransmitter (ACh, at least for some neurons), and so on. Another thing they share is the all or none principle. Have you ever played with dominoes? I don't mean the game, but standing a bunch of them on end, just looking forward to the moment when you tipped the first one over, and the whole chain topples over in series.
You can tilt the first domino only so far and not have it fall over, but once you pass the point of no return, that sucker's goin' down, and all the others will fall down in glorious quick succession. In the same sense, muscle cells and neurons function on an all-or-none principle. Once the stimulus exceeds a certain threshold, the contraction or nerve impulse in that single cell will happen, and it will happen all the way. That principle is fully dependent on those gated ion channels opening, letting Na+ ions flood into the cell.
Action Potential
Despite the differences between the various gated ion channels, they all function on the same basic principle, which involves a cell at rest—remaining polarized as a result of ATP maintaining the active transport of ions. These membranes have a basic resting potential, which has to do with the difference in charge on either side of the membrane. The high concentration of Na+ and Cl- ions on the outside, and the high concentration of K+ ions and negatively charged amino acids on the inside of the neurilemma produce an overall voltage of -70 mV (-70 mV = -0.07 Volts). This means that the inside of the membrane is 70 mV more negative than the outside.
When the membrane potential rises to -55 mV, a threshold is reached, and this point of no return triggers the depolarization of the membrane. The membrane will reach an overall voltage of +30 mV, or 30 mV more positive on the inside of the membrane, before repolarization begins. Depolarization and repolarization form a rapid cycle that lasts about one millisecond, or one thousandth of a second, depending on the type of neuron. Inhibitory neurotransmitters make the membrane potential more than -70 mV, thus making it harder to reach the threshold of -55 mV. This state is called hyperpolarization. The succession of neighboring channels carrying this on creates a nerve impulse, also known as an action potential.
Figure 19.6 illustrates that the voltage-gated Na+ channel has two gates, an activation gate and an inactivation gate; the K+ channel only has one gate. In the Na+ channel the activation gate remains closed when the membrane is maintaining its resting potential. With one gate closed, there is no need to keep the other closed. When the membrane depolarizes, both the activation gate of the Na+ channel, and the single gate of the K+ channel opens (although much more slowly), causing the ions to flow, thus allowing them to reach equilibrium (depolarized).
In order for the membrane to repolarize, the inactivation gate of the gated Na+ channel closes, and the gated K+ channel opens to help restore resting potential. When the membrane is fully restored to its polarized state, the K+ gate closes, the Na+ inactivation gate opens, and the Na+ activation gate closes. During this time the Na+/K+ pump has been working to pump the ions to areas of higher concentration (that pesky active transport again!). Amazing that all of this happens in one millisecond! No wonder an action potential, nerve impulse, happens so fast!
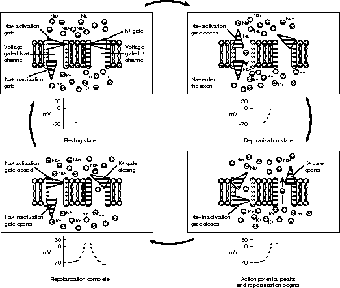
Figure 19.6The gated ion channels involved in an action potential. The Na+/K+ pump, which is used to return the membrane to a polarized state, is not shown here.
Propagation
Remember those voltage-gated channels? The depolarization of one channel causes a change in the voltage of that area of the cell membrane. This causes the neighboring area of the cell membrane to depolarize. This depolarization of the ion channel is followed by an immediate repolarization of that part of the membrane. You might remember from The Structure of the Muscles and Muscle Cells that after a cell is stimulated there is a refractory period, during which a cell cannot be stimulated again. What this means is that the depolarization of one ion channel can trigger the next one to depolarize, but the previous one must recover before it can be stimulated again.
The result of this is that depolarization, and the wave known as the nerve impulse, can travel in only one direction: forward! With the previous ion channel recovering, stimulation cannot travel backward! The ultimate result of this is the need to evolve two sets of nerve fibers, afferent and efferent, to carry messages back and forth.
Once again, propagation from one neighboring ion channel to another is called continuous conduction. This works fine in many circumstances, including the brain, but it is limited by its speed. In the brain the messages don't have far to travel, but in the PNS messages may have to travel 2 meters in a fraction of a second! In this case the neurolemmocytes (Schwann cells) speed up the conduction. The myelin sheath allows the impulse to jump from node to node, producing the much faster saltatory conduction. Just what the PNS ordered!

Excerpted from The Complete Idiot's Guide to Anatomy and Physiology © 2004 by Michael J. Vieira Lazaroff. All rights reserved including the right of reproduction in whole or in part in any form. Used by arrangement with Alpha Books, a member of Penguin Group (USA) Inc.
To order this book direct from the publisher, visit the Penguin USA website or call 1-800-253-6476. You can also purchase this book at Amazon.com and Barnes & Noble.