Cell Theory, Form, and Function: Fluid Mosaic Model of Membrane Structure and Function
Fluid Mosaic Model of Membrane Structure and Function
Membranes have many different functions within a typical cell, such as keeping unwanted viruses out, but probably the most valuable is the partitioning of the cell into functional and segregated compartments. Because of the incredible number and often conflicting biochemical reactions occurring in a cell at any one time, the cell must retain order via structural organization or risk chemical chaos. The internal membranes compartmentalize reactions to prevent interference. The cell membrane also separates life from the nonlife on its exterior. In so doing, an intact and healthy membrane is selectively permeable because it allows substances needed for cell prosperity to enter and attempts to prohibit the penetration of unwanted and unfriendly substances. Unfortunately the system is not always fool-proof. Sometimes unwanted substances pass through the membrane and may cause trouble within the cell.
Bioterms
A glycoprotein is a molecule used as an identification or address for proteins seeking a particular site for bonding. There are many different types of glycoproteins because of the vast array of sugars that may combine with the proteins that compose them.
Interestingly, when a phospholipid is placed in water, it spontaneously folds upon itself to create a double layer, or bilayer. This bilayer phenomenon is also the foundation for the widely upheld fluid mosaic model of membrane structure. The phospholipid molecule has a water-soluble, polar head and two fat-soluble, nonpolar tails. The hydrophobic tails always try to avoid water and face the inside of the bilayer, whereas the hydrophilic head faces the exterior and the interior.
Within the phospholipid bilayer are many different types of embedded proteins and cholesterol molecules whose presence spawned the term mosaic. From scanning electron microscope images, it was observed that the embedded molecules can move sideways throughout the membrane, meaning the membrane is not solid, but more like a fluid. The membranes also have glycoproteins attached to their surface, which aid in their location and identification of food, water, waste, and other membrane traffic. Each cell has a particular glycoprotein structure based on its need to attract or repel membrane traffic. Refer to the illustration Typical membrane, and note the arrangement of the phospholipid molecules.
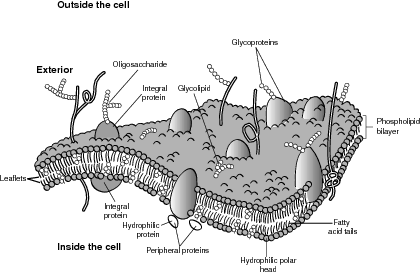
The proteins embedded in the membrane serve many of the membrane functions, such as holding the membrane in a regular, identifiable structure for easy bonding. They also have a specific and unique shape that allows them to function as receptors and receptor sites for attachment to the appropriate raw materials needed for cellular functions. In some cases, the receptor protein is also a signal transducer that begins a series of enzyme-catalyzed reactions to stimulate a particular reaction or function within a cell. Finally, the transport proteins, also called carrier proteins, help substances move across membranes, as described in the next section.
Passive Transport
Passive transport occurs when no energy is required to move a substance, such as water or carbon dioxide, from an area of high concentration to an area of low concentration until the concentration is equal, sometimes across a membrane. The high-to-low concentration gradient is the driving force for passive transport because it fulfills a fundamental law of nature: Things tend to move from a high-energy, ordered structure to a lower-energy, increasing randomness, or increasing entropy state of being. The following are the classes of passive transport:
- Diffusion. This is a good example of how certain molecules, such as oxygen, simply move directly through a membrane in response to the high-to-low concentration gradient. As an example, oxygen diffuses out of the lungs and into the blood for transport to all of the cells.
- Facilitated Diffusion. This is a special type of diffusion that is useful because substances are sometimes too large to move freely through a membrane, or they need to move against a concentration gradient so transport proteins embedded in the membrane assist with the passage. In most cases, the transport protein creates a chemical channel for the passage of a specific substance. Because no energy is expended, the rate of facilitated diffusion depends on the number of transport proteins embedded in the membrane. As an example, glucose is moved by a glucose-transporter protein as it passes through the red blood cell into a body cell.
- Osmosis. This is similar to diffusion except that it refers only to water diffusing through a permeable membrane. Water as a solvent moves from an area of high to low concentration. In biological systems, it is easier to think of water as flowing from a low-solute to a high-solute concentration until the concentration is equal. The solution that has a high-solute concentration is a hypotonic solution relative to another lower-solute concentration or hypertonic solution. Water will continue to osmotically move from the low-solute/high-solvent concentration toward the high-solute/low-solvent concentration until both sides are isotonic, or equal. Osmoregulation is a struggle for all organisms as we continually adjust our cellular water balance for optimal conditions. In your body, the large intestine reabsorbs water by osmosis to help maintain the proper water concentration, which helps to keep your systems from dehydrating.
- Ion channels. These are membrane proteins that allow the passage of ions that would ordinarily be stopped by the lipid bilayer of the membrane. These small passageways are specific for one type of ion, such that a calcium ion could not pass through an iron ion channel. The ion channels also serve as gates because they regulate ion flow in response to two environmental factors: chemical or electrical signals from the cells and membrane movement. This happens in your body when a nervous impulse encounters a gap or synaptic cleft between nerve cells. The electrical stimulation is continued because ion channels are opened to allow specific ions to pass through the receiving membrane, which continues the electrical stimulation to the next nerve cell.
Active Transport
Sometimes substances must be pumped against a concentration gradient, such as the sodium ions (Na+) and potassium ions (K+) pump. So a transport protein and energy, usually adenosine triphosphate (ATP), the energy-rich compound, are needed to push the ions against the gradient. In the case of sodium and potassium ions, maintaining sodium outside and potassium inside the cell is crucial to the functioning of muscles and nerves. The following mechanism illustrates an active transport mechanism:
- Sodium ions inside the cell bind to the transport protein as a phosphate is added from an ATP, which changes the shape of the transport protein.
- The new transport protein structure carries and deposits the sodium to the exterior and bonds with a potassium ion, loses the phosphate group (which again changes the shape of the transport protein), and allows for the return trip.
- The potassium is deposited inside the cell, and a sodium ion and a phosphate are attached to a transport protein to repeat the process.
Endocytosis and exocytosis handle the really big molecules, such as long protein chains or ringed structures, as well as the bulk volume of small molecules. In endocytosis, substances such as food are brought into the cell in a process in which the cell membrane surrounds the particle and moves the particle inside the cell, creating a vacuole or vesicle as a membrane-enclosed container. In exocytosis, waste products or hormones, which are contained in vacuoles or vesicles, exit the cell and their containing membrane is absorbed and added to the cell membrane. There are three types of endocytosis:
- Pinocytosis occurs when the cell absorbs fluid from the exterior, creating a fluid vacuole.
- Receptor-mediated endocytosis is a special type of pinocytosis that is activated by the identification of a receptor protein sensitive to the specific substance.
- Phagocytosis is the engulfing and digesting of substances, usually food, by vacuoles with a lysosome attached (a lysosome is an organelle that contains digestive enzymes).
Endomembrane System
Unlike a prokaryote cell, all eukaryotic cells, regardless of plant, animal, or other origins, are structurally similar and contain mostly the same organelles, with certain exceptions noted. Eukaryotes are compartmentalized by inner membranes to increase active surface area, increase the sophistication of subcellular reactions, and thereby increase overall efficiency.
Within the eukaryotic cell, the endomembrane system is a functional association of membrane-bound organelles that are interconnected or closely connected that build, store, and transfer biomolecules. The biologically important endomembrane organelles are discussed in greater detail in the sections that follow. Refer to the illustration A typical cell.
The nucleus is the centerpiece of the cell. It stores the DNA in the form of chromatin, which is DNA plus a protein, and also serves as the site where RNA copies DNA to begin protein synthesis. The proteins are made in the ribosomes, which are in turn made by the nucleolus, which is also a nuclear component. The nucleolus is where the ribosomal RNA is made and assembled with proteins to make tribosomal subunits. A double nuclear membrane encompasses and separates the nucleus from the cytoplasm. Prokaryotes do not have a well-defined nucleus.
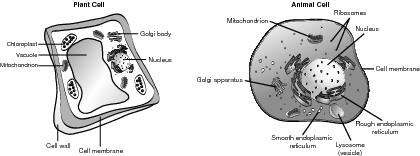
The rough and smooth endoplasmic reticulum is a long, continuous membrane that has many functions within a cell. The rough endoplasmic reticulum (rER) is prominent in cells that create and export proteins because it has ribosomes attached to the membrane. The rER has two important functions, including making more membrane to lengthen the ER and further compartmentalize the cell. Another function is to make secretory proteins, such as antibodies, that are created to function outside of the mother cell. Secretory proteins and other extracellular exports are enveloped by a transport vesicle and are therefore able to move undisturbed through the cytoplasm to the Golgi apparatus.
The rER also connects the nuclear membrane to the smooth ER (sER). The sER builds lipids, regulates the calcium levels so muscles perform correctly, and helps break down toxic substances in the liver. They do not contain ribosomes.
The Golgi apparatus, or Golgi complex, is not connected to the ER, but receives proteins from the ER and modifies them for extracellular export. When a Golgi apparatus receives a transport vesicle from the ER, it sorts the product into like storage areas, chemically marks them for destination points, repackages them in a new transport vesicle, and releases the resulting package to the cell membrane for extracellular export.
The structure of the Golgi apparatus supports its function. Electron microscopy indicates that the structure resembles interconnected, deflated balls or bags. One end serves as a receiving point, the other a shipping center, and the middle contains molecules that sort products and synthesize vesicles to surround them.
Lysosomes are sometimes called the cell's suicide pill because they are single-membrane organelles that contain hydrolytic, digestive enzymes that could easily destroy the cell. Their construction begins in the rER where the enzymes and membranes are joined, and finish in the Golgi apparatus. When fully functional, lysosomes are released and operate mostly in animal cells to perform their four primary functions:
- Subcellular digestion of food particles and nonfunctioning organelles
- Recovery and recycling of certain biomolecules for later use by the cell
- Destruction of harmful foreign particles, such as invading bacteria
- Digestion and removal of the webbing between embryonic fingers
Vacuoles, such as lysosomes, are single-membrane-bound sacs filled with fluid. They also serve four major functions, as explained by their use in the central vacuole found only in plant cells:
- Absorb and store water
- Store enzymes until needed, and metabolic wastes until removed
- Contain attractive pigments to lure pollinators to flowers
- Store toxic chemicals, which also serve as deterrents to herbivores
Energy Production: Chloroplasts and Mitochondria
All green plants have chloroplasts that serve as the location for photosynthesis. Although chloroplasts may be found in all above-ground parts of the plant, most are concentrated in the middle, or mesophyll, of the leaf. Chloroplasts are enclosed in a double membrane that creates a fluid-filled compartment between the membranes, called a stroma. Within the stroma are thylakoids, which are stacked like chips into grana. Within the thylakoid membrane are various types of chlorophyll molecules that capture and convert the energy of light into the chemical energy of chemical bonds. The thylakoid membrane greatly increases the available surface area and houses most of the enzymes and machinery for use in photosynthesis. Each photosynthetic cell contains many chloroplasts, which contain many grana.
Like all subcellular organelles, the function of the mitochondria is related to its structure. The primary purpose of the mitochondria is to conduct cellular respiration, converting the chemical energy of food molecules, such as carbohydrates, into high-energy compounds, such as ATP. Similar to chloroplasts, mitochondria are enclosed by a double membrane that creates a fluid-filled intermembrane space. The second compartment, the mitochondrial matrix, is contained by the highly folded inner membrane. The cristae, or folds of the inner membrane, greatly increase the surface area and contain a multitude of enzymes, so most cellular respiration reactions that produce ATP are completed in the mitochondrial matrix.
Endosymbiosis
An American researcher, Lynn Margulis, proposed in 1966 the hypothesis of endosymbiosis, which may explain the advent of the first eukaryote. According to Margulis, there were two successful invasions of an early anaerobic (one not requiring oxygen) prokaryote, by smaller independent prokaryotes. One of these prokaryote invaders entered the larger prokaryote probably for protection and easy access to nutrients, decided to stay, and began to reproduce independently inside the host cell. Rather than try to evict the invader, the two cells developed a mutually beneficial relationship. The invading cell is thought to be the modern-day mitochondria. A second invasion of similar style, but this time by a photosynthetic bacterium, eventually became a chloroplast. Interesting evidence supports this hypothesis. First, both mitochondria and chloroplasts contain their own DNA, which is separate and different from the rest of the cell. Second, the arrangement of their DNA is circular, a characteristic of prokaryote cells. Finally, both reproduce independently of the rest of the cell.

Excerpted from The Complete Idiot's Guide to Biology © 2004 by Glen E. Moulton, Ed.D.. All rights reserved including the right of reproduction in whole or in part in any form. Used by arrangement with Alpha Books, a member of Penguin Group (USA) Inc.
To order this book direct from the publisher, visit the Penguin USA website or call 1-800-253-6476. You can also purchase this book at Amazon.com and Barnes & Noble.