Inheritance: Meiosis and Sexual Reproduction
Meiosis and Sexual Reproduction
Sexual reproduction is the union of male and female gametes to form a fertilized egg, or zygote. The resulting offspring inherit one half of their traits from each parent. Consequently they are not genetically identical to either parent or siblings, except in the case of identical twins. As hypothesized by Mendel, adults are diploid, signified as 2N, having two alleles available to code for one trait. The gametes must be haploid, signified by N, containing only one allele; so that when two haploid gametes combine, they produce a normal diploid individual. The process where haploid sex cells are created from diploid parents is called meiosis, and it occurs only in the reproductive organs.
Bioterms
Gametogenesis occurs only in the ovaries and testes and represents the formation of haploid egg and sperm as a result of meiosis.
A diploid cell undergoing meiosis first duplicates itself and then divides two times, creating four haploid cells. Meiosis begins with the same G1, S, and G2 stages as mitosis and also ends with a duplicate set of chromosomes. In both processes, the cell divides to form two diploid (2N) offspring; however, meiosis continues with another division, which creates the four haploid gametes, in a process called gametogenesis. In meiosis, several interesting events may happen along the way to provide genetic recombination, an unexpected change in the hereditary genetic material.
Bionote
A chromosome contains numerous genes and is made of two sister chromatids joined by a centromere. The centromere is the region of a chromosome where the two sister chromatids are joined. Homologous chromosomes are inherited from each parent and are the two chromosomes that make up a pair in a diploid cell. They are normally the same length, contain similar genes in the same location, and have a centromere at the same locus.
Mechanism of Meiosis I
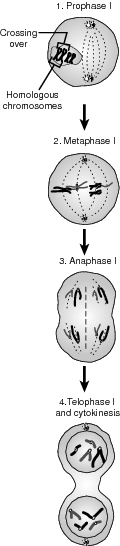
The functional difference between mitosis and meiosis occurs in meiosis I. A synapsis occurs during prophase I, where homologous chromosomes align next to each other. Homologous chromosomes are the matched pair found in a diploid cell. The maternal and paternal homologous chromosomes are made of two sister chromatids that are duplicated copies; so at synapsis, four chromatids are aligned together in a structure called a tetrad, which is a fundamental difference between mitosis and meiosis. In metaphase I, the chromosome tetrads are oriented along the metaphase plate, the equator between the two opposite ends of the cell. In anaphase I, the tetrads split, with the sister chromatids remaining joined at their centromere. When the chromosomes arrive at their respected sides of the cell, in telophase, cytokinesis begins and one cell becomes two. They remain duplicates, but they are still haploid. The two cells now enter meiosis II. Refer to the illustration Meiosis I for a pictorial representation.
Mechanism of Meiosis II
The overall result of meiosis II is to create four haploid sex cells from the two diploid cells that began the process; refer to the illustration Meiosis II for a pictorial representation. Like mitosis, in metaphase II the chromosomes line up along the cell equator, and the paired chromatids separate in anaphase II. This final separation reduces the chromosome number by one-half, creating the haploid sperm and egg. Because of segregational and independent assortment, they may contain completely different alleles.

Abnormalities, Genetic Recombination, Variability
Spontaneous mistakes occur during meiosis that lead to gametes with unusual changes in their genetic structure (makeup). These gene changes lead to an unexpected genetic recombination that, if the organism survives, increases the genetic variability for the population. There are opportunities described next to increase genetic variability during meiosis.
The crossing over of sister chromatids sometimes occurs when they are aligned as tetrads in metaphase I. One chromatid or chromatid piece mistakenly lies on top of a neighboring chromatid. The neighboring nonsister chromatid absorbs the new piece of chromatid into the chromosome and releases the corresponding piece to be absorbed by the first chromosome. The net result is a new genetic recombination, because neighboring chromosomes have exchanged pieces of chromatid that will undergo meiosis as a new component of the chromosome. Refer to the illustration Crossing over.

The random alignment of chromosomes during metaphase I allows equal opportunity for a particular chromosome to migrate into a cell. This type of independent assortment gives rise to exponential gene combinations in the offspring.
Sometimes the spindle fibers fail to separate homologous chromosomes during anaphase I, which overloads one cell with chromosomes and short stocks the other. Likewise, in anaphase II of meiosis II, if a pair of sister chromatids fails to separate and migrates into the same cell, that cell now has too many chromosomes and the other, too few. These scenarios are examples of nondisjunction, which results in the production of gametes with an odd number of chromosomes. Because it often occurs in meiosis, the genetic recombination only affects the X and Y chromosomes, the chromosomes most noted for determining the sex of the offspring, giving rise to the following abnormalities:
- XXy = the offspring is a male with Kleinfelter's syndrome (also includes XXXy, XXXXy, and XXyy; the appearance of a single y chromosome apparently is enough to create a male). Individuals with Kleinfelter's syndrome usually display lanky builds with feminine characteristics such as breast development and poor facial and chest hair growth, and they are mentally retarded and sterile.
- Xyy = the offspring is a normal male, often called a supermale because the presence of an extra y chromosome may contribute to characteristics of increased height, weight, muscular bulk, and aggressiveness
- XXX = The offspring are female, called metafemales or superfemales, and appear normal.
- XO = The offspring are female and have Turner's syndrome (only one chromosome present). Individuals with Turner's syndrome are sterile females that are short in stature, do not sexually mature, and have a thickened web of skin between the shoulders and neck.
Mutations are the primary source of genetic variability because a mutation creates a new gene. Variability is also increased in other ways, such as by the randomness of the union between sperm and egg leading to fertilization.
Mutation
A mutation is a unique type of abnormality and is the greatest source of genetic variability because it creates a change in the nucleotide sequence composing the DNA. Mutations can be either good or bad.

Assuming the daughter cells receive the correct number of chromosomes, problems may arise in the structure of the DNA itself. Mutations involving the rearrangement of the DNA nucleotides is caused in four distinct ways.
A translocation occurs when the DNA double helix is broken and a piece of the chromosome attaches to a neighboring nonhomologous chromosome, making it longer than its homologous chromosome. The donating chromosome is obviously now shorter that its homolog. Refer to the illustration Translocation.

Whenever a segment of a chromosome is broken off and lost, the resulting deletion has serious effects on the transmission of the original genetic material. Refer to the illustration Deletion.

If a deleted segment returns and joins with a homologous chromosome, a duplication of genes has occurred. Refer to the illustration Duplication.

Finally, if a segment breaks loose, reverses, and reattaches in reverse order, an inversion results. Refer to illustration Inversion.
Bionote
A karyotype is a display of an individual's chromosomes that have been stained for easier observation. In humans, the karyotype shows any alteration in the 22 autosomal (genes that code for the body) chromosome pairs or the one pair of sex chromosomes.
As a result of inversions, the genes are still present and the gene number is still the same; in translocations and deletions, however, the resulting mutations may create serious problems, especially in the case of deletions because the “reading” of the genetic code will be altered by an extra omitted gene.
On a smaller scale, a point mutation in a gene is the single exchange of one nucleotide for another. This type of genetic recombination may be too small to affect the overall function of the protein and may not be noticed by the individual, especially if it is an interon which are described in Regulation of Gene Expression in Prokaryotes and Eukaryotes. In other cases, a point mutation may improve the organism by making it more fit, or make it worse by decreasing the fitness thereby lowering its chance of survival.

Excerpted from The Complete Idiot's Guide to Biology © 2004 by Glen E. Moulton, Ed.D.. All rights reserved including the right of reproduction in whole or in part in any form. Used by arrangement with Alpha Books, a member of Penguin Group (USA) Inc.
To order this book direct from the publisher, visit the Penguin USA website or call 1-800-253-6476. You can also purchase this book at Amazon.com and Barnes & Noble.