Anatomy and Physiology: Sliding Home with Sarcomeres
Sliding Home with Sarcomeres
At this point you need to take a good look at the place where the actual contraction happens. A number of fascinating proteins work together to make contraction possible. I will focus on striated muscles (skeletal and cardiac) here, because the proteins are highly organized into parallel stripes (striations) that literally get closer together when the muscle contracts. On a smaller scale, these striations are formed by sarcomeres, which are the unit of muscle contraction.
It is helpful here to take a look at the appearance of a striated muscle cell, for the parallel stripes fall into a clear pattern. Each part of the stripes has a specific name that is identical to the corresponding proteins in the sarcomere. Remember that each striation represents hundred of parallel sarcomeres. In the myofiber as a whole you first see the division into dark A bands, and light I bands, but closer examination reveals a series of other details such as the Z disk, H zone, and M line. Take a moment and try to find these same structures in the diagram of the sarcomere (see Figure 8.6).
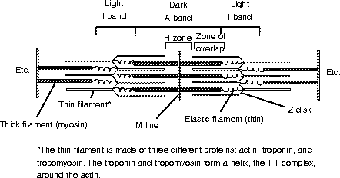
Figure 8.6The sarcomere is the unit of muscle contraction. Compare the parts of the sarcomere to the parts of the previous myofiber diagram. (©Michael J. Vieira Lazaroff)
The biggest details in the sarcomere are probably the thick and thin filaments. The thick filament, as you can imagine, corresponds to the dark A band, and the thin filament corresponds to the light I band. Much of the rest of the sarcomere can also be seen on the myofiber: Z disk, H zone, M line. The only structure that is not visible on the myofiber is the elastic filament (made of a protein called titin), which connects the thick filament to the Z disk.
Shrinking Stripes
When you take a much closer look, you see that there are more details to both the thin and thick filaments, and understanding those details is vital to understanding how a sarcomere works. The thick filament is made of a protein called myosin. Myosin is arranged as a large (thick) filament with numerous tails coming off in all directions, and at the end of each myosin tail is the myosin head. Also on the myosin head is an enzyme called ATPase, which is responsible for converting ATP into ADP, a free phosphate, and energy. The thin filament, however, is far more complex. The principal thin filament protein involved in contraction is called actin, and a helical chain of actin molecules forms the core of the thick filament. On the actin molecule is a myosin binding site, where the myosin head attaches; the attachment of myosin head and actin molecule is called a cross bridge. When left alone, actin and myosin bind together and stay there. A muscle in such a situation would be rigid, which is hardly the state of a relaxed muscle. It is therefore important that the cell have a way to lock and unlock the actin.
Surrounding the actin, locking it as a cable lock locks up a bicycle, is a pair of protein molecules called troponin, and tropomyosin. Because the molecules go together they are referred to as the troponin-tropomyosin complex (or T-T complex for short). The helical T-T complex wraps around the helical actin molecules, with the tropomyosin acting as the cable, and the troponin acting as the lock. When the T-T complex is locked, the tropomyosin covers up the myosin binding sites on the actin, thus making the muscle relaxed, and allowing it to be stretched by other muscles. (I will look at antagonistic pairs in more detail in The Muscles.)
When unlocked, the tropomyosin moves aside, thus exposing the myosin binding sites and allowing the actin-myosin cross bridges to form. The filaments then contract, which means that the thin filaments slide over the thick filaments (hence the sliding filament name) in each sarcomere, thus making the muscle contract (see Figure 8.7). All of this raises two questions: How does the T-T complex get unlocked, and after forming the first cross bridge, how does the muscle avoid staying rigid and actually move? To answer those questions you need to look at two substances: Ca2+ and ATP.

Figure 8.7The shrinking of the space between the Z-discs, due to the sliding of thin filaments over thick filaments, makes muscle contraction possible. (©Michael J. Vieira Lazaroff)
Moving Proteins
The calcium ions act as a key for the troponin, with the keyhole being a place ion the troponin molecule called, conveniently enough, the calcium-binding site. As you should remember, the calcium is released from the S.R. only upon the influx of sodium ions, which was in turn triggered by the ACh stimulus from the motor neuron. At this point, the muscle is free to contract. There is just one problem: the rigid actin-myosin cross-bridge.
To break each cross bridge requires the energy from one ATP molecule. Luckily the ATPase can be found at the end of each myosin head. It is helpful here to understand that forming the cross bridge the myosin head will swivel toward the M line (which acts to anchor the thick filament). By using ATP to break the bond, the swivel action pulls the thin filament (inertia also plays a role here, as an object in motion will stay in motion until acted upon by a force) toward the M line in an action called the power stroke.
By repeatedly forming and breaking the cross bridges, and thus doing repeated power strokes, the thin filaments as a whole will slide closer to the M line, thus making the whole sarcomere shorter. Even though ATP is needed for the active transport that keeps the muscle relaxed, far more ATP is needed to power the repeated power strokes that make contraction possible. All this is summarized in Figure 8.8.

Excerpted from The Complete Idiot's Guide to Anatomy and Physiology © 2004 by Michael J. Vieira Lazaroff. All rights reserved including the right of reproduction in whole or in part in any form. Used by arrangement with Alpha Books, a member of Penguin Group (USA) Inc.
To order this book direct from the publisher, visit the Penguin USA website or call 1-800-253-6476. You can also purchase this book at Amazon.com and Barnes & Noble.